-
メール
yilaibo@shyilaibo.com
-
電話番号
15221734409
-
アドレス
上海市宝山区長江南路180号B区B 650
イレボバイオテクノロジー(上海)有限公司
14例の結腸直腸癌肝転移患者サンプルから治療の新たな標的が発見された――CTSD
日付:2025-11-28読む:0
結腸直腸癌肝転移のジレンマ
結腸直腸癌肝転移は結腸直腸癌zuiによく見られる遠方転移形式であり、約15%から25%の患者は初診時に合併肝転移し、約50%の患者の病歴中に肝転移が発生し、その中の約20%は同時転移(確定診断時に存在)、30%は異時性転移(術後フォローアップ)である。肝臓は結腸直腸癌の血行転移の最も主要な標的器官として、門脈系を通じて転移病巣を形成する。手術による原発性かまどの切除及び転移かまどの切除は依然としてwei一gen治療手段であるが、80%〜90%の肝転移かまどは最初はgen治療性切除ができなかった。
北京大学人民病院胃腸外科の申占龍教授、葉穎江教授と中国ke学院生物物理研究所の李岩教授はこのほど、国際quan威定期刊行物Advanced Scienceで研究論文を発表した。(提供/人民網日本語版・編集/KLM)。(提供/人民網日本語版・編集/KLM)。(提供/人民網日本語版・N-glycosylation modification of CTSD affects liver Metastases in colorectal cancer。同研究では、糖蛋白質組織プロテアーゼD(CTSD)が結腸直腸癌肝転移の標的治療の標的になる可能性があることを指摘した。

N−糖蛋白質群学による結腸直腸肝転移標的の実現
研究者は14例の結腸直腸癌肝転移患者から14対の対の原発巣と転移巣癌組織を分析した:139個のN-グリコシル化蛋白、185個のN-グリコシル化修飾部位、合計490個の完全構造のN-グリコシルペプチドと71個のグリコシルを検出した。差異解析により、組織プロテアーゼD(CTSD)263残基上に

N−グリコシル化修飾はCTSDの折り畳み位置、安定性及び機能活性を変化させる
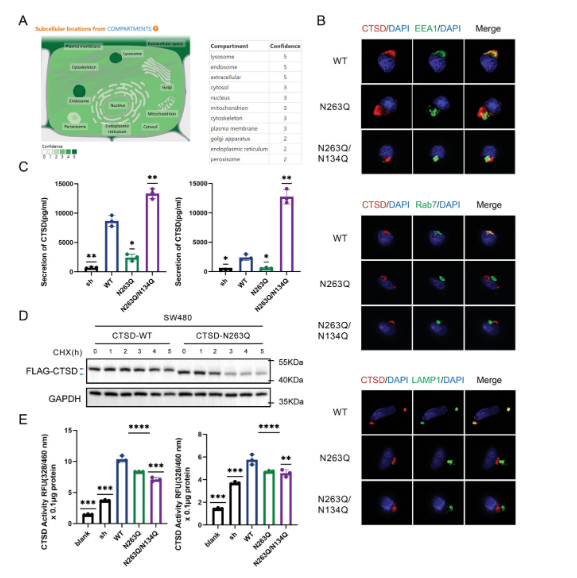
CTSDは、CTSD N 263 QおよびCTSD N 263 Q/N 134 Q細胞において、内体およびリソソームマーカーと共位置決めすることができない(図4 B)。すなわち、CTSD残基263のN−グリコシル化修飾レベルを低下させることは、SW 480細胞コア内体およびリソソームへの輸送を抑制する可能性がある。CTSD WT細胞と比較して、部分的にN−グリコシル化修飾を欠いているCTSD N 263 Q細胞のCTSD分泌量は減少し、一方、N−グリコシル化修飾を欠いているCTSD N 263 Q/N 134 Q細胞のCTSD分泌量は増加した。したがって、CRC細胞は、CTSD残基263のN−グリコシル化修飾が欠落しているため、CTSDを分泌することができない。
また、CTSD WT細胞と比較して、シクロヘキサイミド処理3 h後、CTSD N 263 Q細胞中のCTSDの数は明らかに減少し、CTSDの263残基におけるN−グリコシル化修飾がその安定性に影響を与えたことを説明した。CTSD N 263 Q細胞とCTSD N 263 Q/N 134 Q細胞のCTSD蛍光値はCTSD WT細胞より低かった(図4 E)。具体的には、CTSD残基134及び263のN−グリコシル化修飾レベルを低下させることは、そのプロテアーゼ活性を低下させる可能性がある。以上のように、CTSD残基134及び263のN−グリコシル化修飾は、その折り畳み位置、安定性及び機能活性を変化させる可能性がある。
グリコシル化修飾CTSD下流タンパク質スクリーニング及び同定
CTSD野生型細胞とCTSD N 263 Q細胞の比較蛋白質群解析を行った。研究者はCTSD野生型細胞とCTSD N 263 Q細胞の間に差異発現がある蛋白質を計391個同定した。この分析では、ACADM、PRDX 3、ALDH 2、EHHADH、DHCR 7、TGFB 1 I 1、IDH 2、ALDH 1 A 1の8つの候補分子が生成された。

STT 3 BとDDOSTはCTSDのN−グリコシル転移酵素である
免疫共沈殿によりCTSDと結合したタンパク質が濃縮され、その後質量分析によりこれらのタンパク質が同定される分析の結果、235個の蛋白質とCTSD WTは相互作用し、191個の蛋白質はCTSD N 263 Qと結合した。CTSD WTwanと全相互作用する45個のタンパク質のうち、DDOST(OST 48とも呼ばれる)はwei一のN−グリコシル転移酵素であり、残基263のN−グリコシル化修飾に関与する可能性がある。最初のN−グリコシル転移酵素は、オリゴ糖転移酵素複合体(OST)とも呼ばれ、複数のタンパク質から1つの複合体を構成する。OSTはOST-AとOST-Bの2つの形式で存在し、それぞれSTT 3 AとSTT 3 Bを触媒サブユニットとし、それぞれ初期N-グリコシル化を担当している。触媒サブユニットは独立してN−グリコシル化付加プロセスを完了することができる。しかし、場合によっては、DDOSTのようなさまざまな非触媒サブユニットが必要であり、この過程で基質と安定した錯体を形成するために、私たちはオンライン蛋白質グループ学データベースCPTACを用いて、結腸癌組織と対合正常組織におけるSTT 3 A、STT 3 B、DDOSTの発現レベルを研究し、これら3種類の蛋白質の結腸癌組織における発現の向上を発見した。ターゲットノックアウトSTT 3 A、STT 3 B及びDDOST:STT 3 B及びDDOSTの発現が減少すると、CTSDの分子量は陰性対照に対して著しく低下し、N−グリコシル化から非N−グリコシル化への転換を示した。これらの発見は、CTSD残基263のN−グリコシル化がDDOSTによって制御される可能性があり、STT 3 Bがこの部位の触媒サブユニットである可能性があることを示している。


研究の概要
この研究により、CTSDのN-グリコシル化修飾はその分泌と活性に顕著に影響し、さらにCRC細胞の侵襲/転移能力を変え、肝転移形成に影響することが明らかになった。CTSDグリコシル化は予後/診断バイオマーカーとしての可能性があり、結腸直腸癌肝転移を遮断するために新たな分子介入標的を提供した。
次の記事:SDラットクモ膜下出血モデル